
Retail:
$374.95
$179.99
Savings:
$194.96
 Skip to the content
Skip to the content 
With over 12 million pills sold to more than 320 thousand satisfied customers, Male Extra is one of the leading, most trusted male enhancement products on the market.
100% MONEY BACK GUARANTEE

NO AUTO-SHIP

NO AUTO-BILL

FREE DISCREET SHIPPING
From stronger erections and impressive stamina to improved confidence and even a saved marriage, Male Extra is transforming the sex lives of guys like you every day!
Can’t thank you enough for what Male Extra has done to me in just 1 month. After the first couple of weeks, I noticed my sexual urge skyrocketed. My wife is really impressed with my stamina. Not only did it enhances my manhood, I got better endurance on any physical activity I am doing. This product is really a marriage/life saver!!!

I admit it after 10 days I thought: “another scam” because I did not notice any improvement. But after 20 days my penis was getting firmer and after 1 month no doubt, I found my erection is satisfactory. Today it’s been almost 2 months that I use Male extra daily and you can not know the happiness of waking up with a good old morning erection lol, I recommend Male Extra because it really works

Taking MaleExtra every day. It gives you hard erections and it also gives you the drive to go that extra mile of satisfaction. Whenever I take MaleExtra the results are just impeccable. I’m a very happy man and will keep on using it. This isn’t no scam this is the real thing – especially without any side effects as well!

From the 2nd and 3rd week I felt the effects – harder erection and my libido increased – I could make love 3 times per day. I did a month with male extra and it works. Thanks to the male extra team!

I purchased it mainly to improve my sexual life as I felt recently my need for that. Since 15 days approximately I started to feel the difference as I started to feel my sex drive ready all the time.
Male Extra uses a powerful fusion of proven, safe and natural ingredients to help increase the size, hardness and sustainability of your erections and improve your sexual performance.
It does this primarily by increasing the natural nitric oxide levels in your blood. Nitric oxide is a vasodilator, which means it relaxes and widens your blood vessels, improving the flow of blood and oxygen through your body.
By improving blood flow to your penis, Male Extra gives you a solid erection that stays harder for longer, and more intense, pleasurable orgasms.
Improved blood flow also enables more oxygen and nutrients to be delivered more quickly to your cells, delaying fatigue during physical activity. Which means you’ll have the stamina to perform better and last longer in bed.


During an erection, the tissues in your penis fill with blood. The more blood these tissues hold, the bigger and harder your penis becomes

Male Extra expands the blood vessels leading to the erectile tissues in your penis, which means more blood will flow into your penis when you have an erection


This increase in blood flow gives you notably bigger, thicker and harder erections that last longer and means you’ll experience stronger, more intense orgasms.

Bigger, Harder Erections – with reliable, solid erections that stay harder for longer, both you and partner will really notice the difference

Powerful Orgasms – with increased blood flow giving you stronger erections you’ll also experience more intense and pleasurable orgasms

Heightened Sex Drive – increase your sexual desire and boost your your libido, making you in the mood for sex and ready for action

Improved Performance – Male Extra gives you the stamina and the staying power you need to make an impression in the bedroom

Increased Confidence – take back control in the bedroom, confident in your ability to give your partner what they really want
Male Extra is the only male enhancement product to use pomegranate ellagic 40% and L-arginine HCL in our formula – you won’t find these super powerful, erection-boosting, libido-enhancing ingredients in any other male enhancement product.
These ingredients have been specifically chosen for their proven ability to significantly improve erection quality and sexual performance. They are backed by multiple studies proving their effectiveness.
In fact, FIVE out of the seven ingredients in our formula are unique to Male Extra! That means you will not find them in ANY other formula on the market.
Every single ingredient in Male Extra is 100% natural and safe. Which means you get proven results with NO side effects.

Ellagic acid is a polyphenol antioxidant found in fruits and vegetables, and in particularly high amounts in pomegranate. A number of studies have demonstrated its effectiveness at increasing blood flow to the penis, improving erection quality and improving sexual performance.
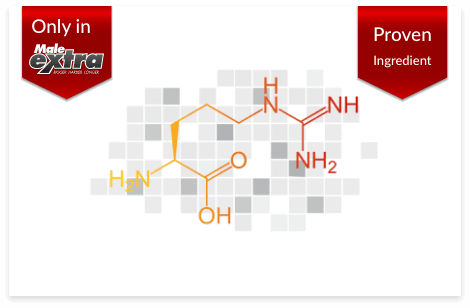
Numerous studies have demonstrated l-arginine’s effectiveness on erection quality and male sexual performance. Your body uses this amino acid to produce nitric oxide, which improves blood flow to your penis, giving you bigger, harder erections.

Used in Chinese medicine for thousands of years as a natural aphrodisiac, many studies have proven the powerful ability of this type of fungus to increase sex drive and sexual function.
Scientists believe this is due to two chemicals in cordyceps called deoxyadenosine and cordycepic acid, which appear to have a direct effect on the area of the brain responsible for controlling sexual desire.

This essential mineral plays a vital role in testosterone production. Low testosterone levels can cause havoc with your libido and your erections, yet most men don’t get enough zinc in their diet. Which means it’s likely you’re not getting enough of it either.
The zinc in Male Extra helps protect you against zinc deficiency, keeping your testosterone at healthy levels and your libido firing on all cylinders.
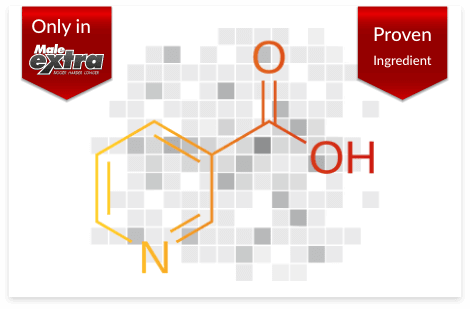
Also known as vitamin B3, studies have shown that niacin can improve erection quality and longevity by relaxing and expanding the blood vessels in the penis, boosting the amount of blood that flows to it during an erection.
Niacin also helps reduce tiredness and fatigue, which means you’ll experience more energy and stamina in the bedroom.

MSM is an organic form of sulphur, a mineral that, as well as supporting blood flow, is crucial to your cell and tissue health – including those in your penis. Without enough MSM, your cells can become weak and your body cannot build strong and healthy new cells.
The sulfur-rich MSM in Male Extra stimulates the growth of healthy new, flexible cells in your penis that can hold more blood during an erection, helping you achieve bigger, firmer erections.
An essential amino acid, L-methionine is thought to work by blocking the conversion of histidine into the hormone histamine.
Histamine plays a key role in achieving climax; the higher the levels of it, the more quickly you are likely to ejaculate, which is why high levels of histamine have been linked to premature ejaculation.
By lowering your histamine levels, L-methionine can help delay ejaculation, helping you last longer in bed.
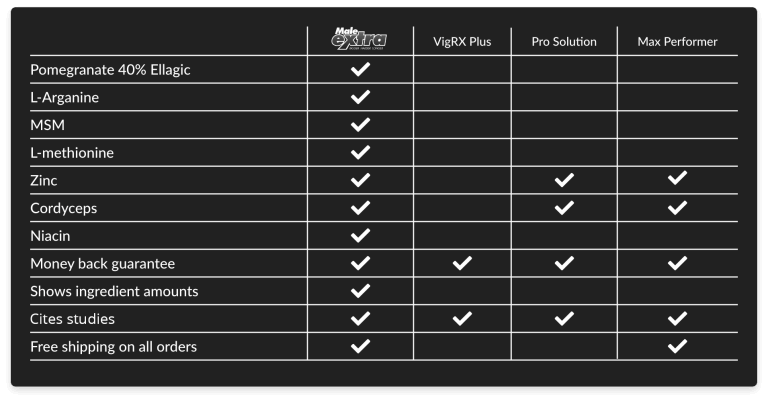
See for yourself how Male Extra stacks up against other male enhancement products on the market.
Just take 3 Male Extra capsules daily with a meal. The powerful, proven ingredients will build up in your system over a period of 30+ days.
During this time you will notice stronger, harder, longer-lasting erections, more intense orgasms and improved sexual stamina.
You will only benefit from these effects as long as you keep taking Male Extra, otherwise the ingredients will slowly disappear from your system. In order to continue to see results, it is important you continue to take Male Extra every day.
Male Extra erection gel delivers powerful erection-enhancing ingredients right where you need it, when you need it.
Just rub directly onto your penis before sex. The active ingredients quickly stimulate blood flow to give you fast and powerful erections.
Order 3 bottles of Male Extra and we’ll send you 1 tube of erection gel absolutely FREE!
Order 5 bottles of Male Extra and we’ll send you 2 tubes of erection gel absolutely FREE!


We’re so confident you’ll experience your best sex ever with Male Extra that we offer you an industry-leading, 100 day money back guarantee. That’s longer than any other male enhancement product on the market! But if, after using MaleExtra as directed for at least 50 days, you don’t believe it has helped improve your sexual performance, simply email us within 50-100 days of receiving your order and we will refund your money, excluding a $15 fixed fee to cover shipping and admin costs*.
*Terms and conditions apply. Guarantee applies to purchases of over one month’s supply and only on first purchase. Click here to read the full terms of our refund policy.
Safe and Natural, Proven Ingredients
Enjoy Bigger, Stronger, Harder Erections
Increase Blood Flow To Your Penis
Be a Winner In The Bedroom
Proven ingredients for bigger, harder erections, better orgasms and improved stamina. Guaranteed.

Discreet Billing and Shipping

256bit Security On All Orders

NO auto billing




Just take three capsules every day with your breakfast or another meal, and enjoy bigger, harder erections and a super-charged sex life! You will only benefit from these effects as long as you keep taking the product, otherwise the ingredients will slowly disappear from your system. In order to continue to see results, it is important you continue to use the product every day.
Results vary between different people, but most of our customers start seeing results within the first few weeks of taking Male Extra. The ingredients will build up in your system over a period of 30-60 days. During this time you will notice stronger, harder, longer-lasting erections, more intense orgasms and improved sexual stamina. You will only benefit from these effects as long as you keep taking Male Extra, otherwise the ingredients will slowly disappear from your system. In order to continue to see results, it is important you continue to take it every day.
The ingredients are designed to increase blood flow to your penis, giving you bigger, harder, longer lasting erections, as well as boosting your sex drive. You will only benefit from these effects as long as you keep taking Male Extra.
No. Male Extra is NOT a permanent penis enlargement solution. Male Extra increases the amount of blood that your penis receives during an erection. This means you will see a notable difference to your erect penis size only. These results will last only as long as you keep taking the product.
Each bottle of Male Extra contains 90 capsules. Each serving size is 3 capsules. This means one bottle will last you one month if you take it every day as directed.
Male Extra contains L-arginine, Pomegranate 40% ellagic acid, MSM (methyl sulfonyl methane), L-Methionine, Zinc, Cordyceps and Niacin (vitamin B3). All are safe and natural ingredients with many studies and trials documenting their effectiveness.
Although Male Extra is formulated from safe and natural ingredients, you should always check with your doctor or health practitioner if you are currently taking any medication to see whether this supplement is suitable for you.
Yes. Men of all ages can use Male Extra. Many of the studies proving the effectiveness of it’s ingredients in improving sexual function were carried out on senior citizens. However, if you are currently taking any medication or have any existing health issues you should check with your doctor or health practitioner before taking this supplement.
All orders are packaged and on their way to you within 24-48 hours. Once dispatched, here is when you can expect to receive your order:
USA: 3-7 working days
UK: 3-7 working days
Europe: 3-20 working days
Rest of the world: 5-21 working days
Please note that COVID-19 restrictions in some countries may cause delays to your delivery. This is due to special measures put in place to ensure the safety of our suppliers and delivery teams. We apologise for any inconvenience this may cause and thank you for your continued patience. If you have any questions, please do not hesitate to contact us.
Your order will be shipped in plain, discreet packaging. You will be emailed a tracking number once your order has been dispatched.You can track your order by clicking here.
Your privacy is of utmost importance to us. Your order will be packaged in completely discreet packaging – no one will know what you have ordered. The name Male Extra won’t show on your credit card statement either – you will be billed from “Payments.mn”.
Our money back guarantee is industry leading! You can purchase Male Extra with confidence, knowing that it’s backed with a no risk, no hassle, 100 day money back guarantee. That’s longer than any other male enhancement product on the market. If, after using MaleExtra as directed for at least 50 days, you don’t believe it has helped improve your sexual performance, simply email us within 50-100 days of receiving your order and we will refund your money, excluding a $15 fixed fee to cover shipping and admin costs. Please note that terms and conditions apply.The guarantee applies to purchases of over one month’s supply and only on first purchase. Click here to read the full terms of our refund policy.
We are a well established company who have dedicated the last 15 years to research and development of innovative health products. Our dedicated customer service team is available 24/7 to answer any questions you have about any of our products.
*These statements have not been evaluated by the Food and Drug Administration. These products are not intended to diagnose, treat, cure, or prevent any disease. These products do not require a prescription or doctor approval as they are herbal and do not contain any prescription ingredients. Results in the Testimonials may not be typical and your results may vary. Not all people will obtain the same results. These products are intended to be used in conjunction with a healthy diet and regular exercise. Consult your physician before starting any diet, exercise program or taking any diet pill to avoid any health issues.
We use mouth masks when packing your order.
We use medical grade hand sanitizer regularly and gloves when packing.
Our delivery services use no-contact drop-off.
